
- Štítky blogu
Určení mechanismu rezistence u jedinců Brassicogethes aeneus F. prostřednictvím molekulárních analýz
Ludvíková M., Seidenglanz M.
Agritec Plant Research s.r.o., Zemědělská 16, 78701 Šumperk
Úvod
V letech 2021–2024 a během prvních 9 měsíců roku 2025 byla u populací blýskáčka řepkového (Brassicogethes aeneus F.) pocházejících z různých lokalit České republiky testována molekulárními metodami přítomnost mutací L1014F a M918T, způsobujících rezistenci vůči pyretroidům. Běžnými laboratorními testy (Adult vial tests; International Resistance Action Committee), které jsou založeny na sledování reakcí dospělců kontaktně vystavených vzrůstajícímu spektru dávek ú.l. pyretroidů, byli z jednotlivých populací k molekulárním analýzám vybráni jedinci, kteří přežili 24 hodin trvající expozici registrované nebo vyšším dávkám. Z těchto fenotypově rezistentních jedinců byla izolována DNA, provedena PCR amplifikace oblasti domény II sodíkového kanálku, sekvence byly purifikovány a odeslány na sekvenování. Následně byla vyhodnocena přítomnost mutace L1014F nebo M918T. Celkem bylo takto testováno 494 jedinců fenotypově rezistentních k pyretroidům na přítomnost mutace L1014F (kdr) a 131 jedinců na přítomnost mutace M918T (s-kdr).
V případě škůdců řepky (a polních plodin obecně) a jejich snížené citlivosti (rezistence) k pyretroidům je možné mluvit o dvou hlavních typech mechanismů rezistence. První je založen na modifikaci vazebného místa pro pyretroidní molekulu, druhý je způsoben nárůstem metabolické detoxifikace účinných látek (Vontas et al., 2010). Mutace, které se projevují sníženou citlivostí sodíkové pumpy k pyretroidům se z hlediska významu a síly dopadu dělí na kdr (knock down resistance) a super kdr (s-kdr). Metabolická rezistence je převládající mechanismus rezistence u českých a obecně i evropských populací blýskáčků. Do tohoto procesu jsou zapojeny enzymy ze skupiny oxygenáz, monooxygenázy Cytochromu P450, popř. i jiné skupiny enzymů (Zimmer et al., 2014).
Cíl
Cílem práce bylo u populací blýskáčka řepkového (Meligethes aeneus F.) pocházejících z různých lokalit ČR testovat molekulárními metodami přítomnost mutací L1014F a M918T, způsobujících rezistenci vůči pyretroidům.
Metody
Běžnými laboratorními testy (Adult vial tests; International Resistance Action Committee), byl proveden výběr rezistentních jedinců z jednotlivých populací blýskáčka řepkového. Pro molekulární analýzu byli použiti jedinci, kteří se v IRAC testech projevili jako fenotypově rezistentní k pyretroidům. Šlo o jedince, kteří bez výrazných projevů postižení přežili kontaktní expozici (24 hodin; světlo: tma = 12: 12 hodin; 18–20 °C) vysokým dávkám těchto pyretroidů: lambda-cyhalothrin (pro blýskáčky od dávky 7.5 g a.i./ha); cypermethrin (pro blýskáčky od dávky 5 g a.i./ha) a tau-fluvalinate (pro blýskáčky od dávky 48 g a.i./ha) a v roce 2025 zařazen také etofenprox (pro blýskáčky od dávky 57.5 g a.i./ha).
Rezistentní jedinci byli po skončení IRAC testu zamraženi a následně byli analyzováni na přítomnost mutace udělující kdr a s-kdr typ rezistence. Jednotliví brouci byli homogenizováni v kapalném dusíku a pomocí Quickgene DNA tissue kit (Kurabo Industries, Německo) z nich byla izolována genomová DNA dle protokolu výrobce. Oblast domény II sodíkového kanálku v oblasti mutace s-kdr byla amplifikována prostřednictvím metody PCR na základě metodiky dle Nauen et al. (2012); použity byly primery PB1, PB2, PB3 a PB4, složení reakční směsi a teplotní podmínky dle uvedené publikace. Pro testování oblasti kdr byly využity primery nově navržené na základě sekvence dostupné v GenBank: AF354457.1.
Pro amplifikaci oblasti kdr bylo složení reakční směsi pro PCR: 20–50 ng DNA, 1x Dream Taq PCR buffer, 100 μM každého nukleotidu, 0,4 μM každého primeru a 1 U Dream Taq polymerázy (Thermo Scientific). Teplotní profil PCR reakcí byl 94 °C /10 min + 35x (94 °C /30 s, 55 °C /30 min, 72 °C /60 s) se závěrečnou elongací 72 °C /5 min. 10 μl vzorku po PCR reakci bylo pro kontrolu naneseno na 1,5% agarózový gel barvený ethidium bromidem, nasvíceno UV světlem a nasnímáno.
Amplifikované sekvence oblastí mutací kdr a s-kdr byly následně purifikovány Isolate II PCR and gel kit (Meridian Bioscience) a odeslány na sekvenování. Získané sekvence byly analyzovány v programu Sequence Scanner (Applied Biosystems, Foster City, CA). Na základě sekvencí byl vyhodnocen genotyp jednotlivých brouků (SS/SR/RR – senzitivní versus rezistentní genotyp).
Výsledky
Při použití primerů PBtest F+R byla výsledná délka amplifikovaného produktu 233 bp, při použití primerů a metodiky dle publikace Nauen et al. (2012), byla výsledná délka amplifikovaných PCR produktů 510 bp.
Mutace M918T (s-kdr) nebyla během testování detekována u žádného vzorku. Mezi výše uvedenými testovanými vzorky byl během molekulárních analýz v r. 2021 objeven 1 vzorek s heterozygotním (T/C) genotypem v oblasti kdr mutace L1014F– výsledek jeho sekvenování je uveden na obrázku.
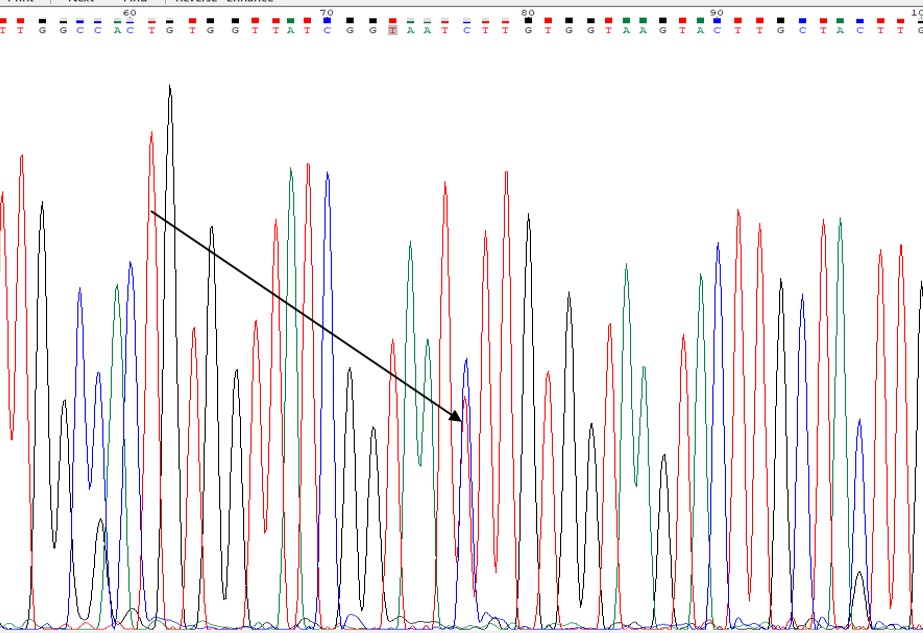
Obrázek 1. Sekvenování heterozygot
Jednalo se o vzorek, který přežil expozici tau-fluvalinate 48 g a.i./ha, pocházející z lokality poblíž Českých Budějovic. Jde o první potvrzení tohoto typu rezistence nejen u populací blýskáčků v ČR, ale i ve střední Evropě.
U tohoto vzorku bylo tedy na území ČR poprvé prokázáno, že jde o rezistenci vyvolanou mutací, projevující se obměnou aminokyselin v proteinu, který je součástí vazného místa (receptoru) na nervovém vlákně (místo, kde se pyretroidní účinná látka za normální situace váže) a ne pouze o metabolickou rezistenci související s nárůstem aktivity některých enzymů (předpokládá se, že cytochromu P450 – monooxygenáz; popř. jiných skupin enzymů) v tělech rezistentních jedinců.
Navzdory počtu sekvenovaných vzorků nebyl v dalších letech v testovaném souboru objeven žádný vzorek nesoucí kdr mutaci L1014F. Molekulární analýzy tedy prokázaly, že u českých populací jsou přítomné oba výše zmíněné typy rezistence (jak metabolická, tak rezistence díky mutaci L1014F), nicméně metabolický typ rezistence zcela převládá a má výrazně vyšší praktický dopad.
Poděkování
Výsledek vznikl za podpory Ministerstva zemědělství, grant QK21010332.